
Comment: Pair-bonding makes drug use less appealing via dopamine receptor mechanism.
Abstract
Hoewol de beskermjende effekten fan sosjale bondels op drugsgebruik / misbrûk binne goed dokumintearre, wy witte net wat oer de ûnderlizzende neurene meganismen. It gebrûk fan de prairie vole (Microtus ochrogaster)—a socially monogamous rodent that forms long-term pair bonds after mating—we demonstrate that amphetamine (AMPH) conditioning induced a conditioned place preference (CPP) in sexually naive (SN), but not pair-bonded (PB), males. Although AMPH treatment induced a similar magnitude of dopamine release in the nucleus accumbens (NAcc) of SN and PB males, it had differential effects on NAcc D1 receptor (D1R) binding. Specifically, AMPH treatment increased D1R binding in SN, but decreased D1R binding in PB males. NAcc D1R, but not D2 receptor, antagonism blocked AMPH-induced CPP in SN males and NAcc D1R activation before AMPH conditioning enabled AMPH-induced CPP in PB males. Together, our data demonstrate that pair-bonding experience decreases the rewarding properties of AMPH through a D1R-mediated mechanism.
Ynlieding
It is goed erkend dat drugsgebrûk en misbrûk djippe gefolgen hawwe op in ferskaat fan sosjale gedrach, lykas sosjale ferbining (Jonge et al., 2011a). Reciprocally, social factors, such as the presence or absence of strong social attachments during development or throughout adulthood, may impact drug use and the vulnerability to drug abuse. For example, strong adolescent–parent attachments have been associated with decreased odds of substance use problems (Ellickson et al., 1999; Bell et al., 2000). Dêrnjonken binne yn yndividuen dy't al ferslavend binne, hokke relaasjes tusken spousels helpe by it opnimmen fan drugsbedriuw (Kosten et al., 1987). Yn tsjinstelling dan kinne swakke maatskiplike oanfetsjes substansje misbrûk meitsje fan kwetsberens as ûnfeilige adult attachmentstilen wurde positiv ferbûn mei alkohol en yllegaal substans misbrûk (Brennan en Shaver, 1995; Vungkhanching et al., 2004; Caspers et al., 2005).
The neural mechanisms underlying the interaction between social bonding and drug use/abuse are still largely unknown; however, the mesolimbic dopamine (DA) system, particularly DA neurotransmission within the nucleus accumbens (NAcc), may be involved (Jonge et al., 2011a). Studies yn 'e prairiefolle (Microtus ochrogaster) - in sosjaal monogamous rodent dat duorsume bondels biedt tusken folwoeksenen (dû, paadbondingen) (Insel en Hulihan, 1995; Mattson et al., 2001) - sjen litte dat NAcc DA sawol de formaasje en ûnderhâld fan paadbondingen (Gingrich et al., 2000; Aragona et al., 2003, 2006). Fierder is NAcc DA neurotransmission aktyf fan alle bekende drugs fan misbrûk (Di Chiara en Imperato, 1988; Bergman et al., 1990; Koob en Nestler, 1997; Nestler, 2005) en repetitive psychostimulante medisyngenoaten DA frijlitting en resepsor-sensibiliteit lykas de morphology fan NAcc neurons (Henry et al., 1989; Robinson en Kolb, 1999). Dizze wizigingen binne tocht oan drugs-induzearre modifikaasjes yn gedrach (Robinson en Becker, 1986), ûnder oaren sosjale gedrach (Fiorino en Phillips, 1999).
As both pair-bonding behavior and its regulation by DA neurotransmission within the NAcc have been well characterized in the prairie vole (Gingrich et al., 2000; Aragona et al., 2003, 2006), recent efforts have been made to establish this species as an animal model in which to examine the behavioral interaction between social bonding and drug use/abuse, as well as its underlying neural mechanisms. Amphetamine (AMPH) has been shown to have rewarding properties in prairie voles, as AMPH conditioning induces a conditioned place preference (CPP) in both sexes (Aragona et al., 2007; Liu et al., 2010; Jonge et al., 2011b); this process is mediated by DA neurotransmission in the NAcc (Curtis en Wang, 2007; Liu et al., 2010). Most interestingly, repeated AMPH exposure inhibits the formation of mating-induced partner preferences in male prairie voles and this AMPH-induced impairment of pair bonding is regulated, at least in part, by NAcc DA D1-type receptor (D1R) activation (Liu et al., 2010). In the present study, we investigated the effects of pair-bonding experience on AMPH-induced CPP. We hypothesized that pair-bonding experience would decrease the rewarding effects of AMPH and that NAcc DA may be involved in this behavioral phenomenon.
Materialen en metoades
SUBJECTS.
Subjekten wiene manlju prairievels (M. ochrogaster) from a laboratory breeding colony. Subjects were weaned at 21 d of age and housed in same-sex sibling pairs in plastic cages (12 × 28 × 16 cm). Water and food were provided ad libitum. All cages were maintained under a 14:10 light:dark cycle and the temperature was kept at 20°C. Subjects ∼75 d of age were either continuously housed with their same-sex sibling [and thus kept sexually naive (SN)] or paired with an unrelated intact female for 2 weeks to become pair bonded (PB). Both SN and PB subjects were tested at ∼90 d of age.
AMPH CONDITIONING AND CPP TESTING.
Dizze prosedueres waarden dien as earder beskreaun (Liu et al., 2010; Jonge et al., 2011b). Briefly, the testing apparatus for CPP consisted of two cages (12 × 28 × 16 cm), one black with a metal top and one white with a mesh top, joined by a hollow tube (7.5 × 16 cm). Although prairie voles generally tend to prefer the white over the dark cage (Aragona et al., 2007), there are a great deal of individual differences in this preference. Therefore, on day 1, we tested all subjects for their initial cage preferences during a 30 min pretest. During this test, all subjects were allowed free access to both cages and we quantified the amount of time each individual spent in each cage. On days 2–4, subjects received two 40 min conditioning sessions 6 h apart. In the morning session, subjects received either 1.0 mg/kg AMPH (Sigma) dissolved in 0.9% saline (SN-AMPH and PB-AMPH groups) or saline alone (SN-saline and PB-saline groups) and were placed in the cage in which they had spent less time during the pretest (conditioned cage). In the afternoon session, all subjects received an injection of saline and were placed in the other cage. On day 5, subjects were tested again for cage preferences in a 30 min posttest. Immediately after the posttest, subjects were rapidly decapitated and their brains were frozen on dry ice. Brain sections were subsequently processed for D1R and DA D2-type receptor (D2R) autoradiographic binding.
BRAIN MICRODIALYSIS AND HPLC-ELECTROCHEMICAL DETECTION ANALYSIS.
Mikrodialysis problemen waarden neamd beskreaun (Curtis en Wang, 2007) and were implanted into the left NAcc (stereotaxic coordinates from bregma: anterior 2.1 mm, lateral 0.6 mm, ventral 6.3 mm) under sodium pentabarbitol anesthesia (1 mg/10 kg body weight). Animals were allowed to recover overnight and were then tested the following morning. Probes were perfused continuously at 2.3 μl/min with a solution isotonic for sodium (144 mm NaCl), potassium (2.8 mmKCl), calcium (1.2 mm CaCl2), and magnesium (0.9 mm MgCl2) (Sved en Curtis, 1993).
Following overnight recovery, four 20 min baseline samples were collected into vials containing 5 μl of 0.1N perchloric acid. Thereafter, subjects received an intraperitoneal injection of AMPH (1.0 mg/kg) and dialysate samples were continuously collected at 20 min intervals for 3 h. Dialysate samples were immediately frozen at −80°C until analyzed. The amounts of DA and DOPAC in each sample were determined using HPLC with electrochemical detection (ECD), as described previously (Curtis en Wang, 2007). At the end of the sampling period, subjects were killed to assess probe placement.
DA RECEPTOR AUTORADIOGRAPHY.
Coronal brain sections (20 μm) at 120 μm intervals were processed for DA receptor autoradiographic binding using an established method (Aragona et al., 2006). Briefly, sections were rinsed in 50 mm Tris-HCl, pH 7.4, and incubated in 50 mm Tris-HCl ion buffer containing 120 mm NaCl, 5m KCl, 2 mm CaCl2, en 1 mm MgCl2 mei [125I]SCH 23982 (D1R ligand) or [125I]2′-iodospiperone (D2R ligand) (PerkinElmer). Thereafter, sections were fixed in 0.1% paraformaldehyde and rinsed thoroughly in Tris-HCl ion buffer. Slides were dipped in distilled water, blown dry, and exposed to BioMax MR film (Kodak) to generate autoradiograms. The optical densities of D1R and D2R binding in the NAcc and caudate putamen (CP) were quantified in three anatomically matched brain sections per animal from autoradiograms using a computerized image program (NIH Image 1.64).
STEREOTAXIC CANNULATION AND MICROINJECTION.
Subjects were anesthetized with sodium pentobarbital and 26-gauge bilateral stainless steel cannulae (Plastics One) were stereotaxically implanted and aimed at the NAcc, as described previously (Aragona et al., 2003). Subjects were allowed to recover for 3–7 d. On each of the 3 d of conditioning, 30 min before AMPH injections, subjects received microinjections of either artificial CSF (200 nl/side) or CSF containing the D1R agonist SKF 38393, the D1R antagonist SCH 23390, or the D2R antagonist eticlopride (Sigma). After CPP testing, all subjects were rapidly decapitated and their brains were extracted to verify injection sites histologically. Subjects with misplaced cannulae were excluded from data analysis.
DATA QUANTIFICATION AND STATISTICAL ANALYSIS.
CPP waard bepaald troch in paar-samples t test comparing the amounts of time that subjects spent in the conditioned cage between pretests and posttests. Cage entries between pretests and posttests were also analyzed by a t test to assess whether AMPH, a D1R agonist, D1R, or a D2R antagonist affected locomotor activity. Absolute amounts of baseline DA and DOPAC in dialysates were compared between groups using a t test. For assessment of AMPH effects across time, the amounts of DA and DOPAC in each baseline and post-AMPH sample were expressed as a percentage of the mean baseline amount. These values then were analyzed by repeated-measures ANOVA followed by a Student-Neuman–Keuls (SNK) post hoc test. Finally, group differences in the densities of D1R and D2R binding in the NAcc and CP were analyzed by a two-way ANOVA followed by a SNK post hoc toets.
EXPERIMENTAL DESIGN.
Experiment 1 was designed to reveal the effects of pair-bonding experience on AMPH-induced CPP. SN and PB males were pretested in the CPP apparatus. They were then divided into four groups that received injections of saline (n = 5 for SN males and n = 9 for PB males) or AMPH (1.0 mg/kg; n = 8 for SN males and n = 8 for PB males) during morning conditioning sessions over the next 3 d (Liu et al., 2010). Thereafter, all subjects received a CPP posttest.
Experiment 2 compared AMPH-induced DA release in the NAcc between SN (n = 6) and PB (n = 5) males. Subjects were implanted with a microdialysis probe aimed at the NAcc. After overnight recovery with continuous perfusion of an isotonic solution through the probes, four 20 min baseline dialysate samples were collected. Thereafter, subjects received an intraperitoneal injection of AMPH (1.0 mg/kg) and dialysate samples were continuously collected every 20 min for 3 h. These samples were subsequently analyzed for DA and DOPAC concentrations using HPLC-ECD analysis (Curtis en Wang, 2007).
Experiment 3 examined the effect of interactions between pair bonding and AMPH treatment on DA receptor binding in the NAcc. Brain sections from the subjects tested in experiment 1 were processed for D1R and D2R binding using receptor autoradiography.
Experiment 4 tested the role of NAcc DA receptors in AMPH-induced CPP. SN males were implanted with guide cannulae bilaterally aimed at the NAcc. After 3 d of recovery, subjects received a CPP pretest and were then randomly assigned into one of five experimental groups [intra-NAcc injections of CSF (n = 8) or intra-NAcc injections of CSF containing a low (4 ng/side; n= 8) or high (100 ng/side; n = 6) dose of a D1R antagonist, SCH 23390, or a low (4 ng/side; n = 8) or high (100 ng/side; n = 7) dose of a D2R antagonist, eticlopride]. Thirty minutes later, subjects received an injection of AMPH (1.0 mg/kg, i.p.). This procedure was repeated for 3 consecutive days during AMPH conditioning. Thereafter, subjects received a CPP posttest.
Experiment 5 examined the role of NAcc D1Rs in mediating AMPH-induced CPP in PB males. PB subjects were divided into three groups, receiving intra-NAcc injections of CSF (n = 10) or CSF containing a D1R agonist, SKF 38393 (0.4 ng/side; n = 12), or a D1R antagonist, SCH 23390 (4 ng/side; n = 10), before AMPH conditioning. Brain cannulation, AMPH injections, and CPP tests were the same as described in experiment 4.
results
Pair-bonding experience decreases the rewarding properties of AMPH
In our previous study, AMPH treatment impaired mating-induced partner preferences in male prairie voles, indicating an inhibitory effect of AMPH exposure on pair-bonding behavior (Liu et al., 2010). Yn 'e hjoeddeiske stúdzje testten wy de omkearde relaasje: de effekten fan pear ferbiningûnderfining op AMPH-belje. Trije dagen fan kondysje mei 1.0 mg / kg AMPH feroarsake in CPP yn SN-manlju (t = 2.45, p <0.05) mar net by manlju dy't 2 wiken mei in wyfke wiene (ie, PB-manlju) (Fig. 1a). Saline injections had no effect in either group. Importantly, no differences were found in the frequencies of the animals’ cage crossings between pretests and posttests, suggesting that impaired CPP in PB males was not due to altered locomotor activity during the behavioral test (Fig. 1b).

- Download figuer
- Iepenje yn nije ljepper
- Download powerpoint
{kind=link}
Figure 1.
AMPH conditioning induces a CPP in SN but not PB male prairie voles. a, SN or PB males that received saline (SN-saline or PB-saline, respectively) during 3 d of conditioning did not form a CPP, i.e., they spent equal amounts of time in the conditioned cage during the 30 min pretests and posttests. AMPH conditioning induced a CPP in SN males (SN-AMPH), but not in PB males (PB-AMPH), indicating that pair-bonding experience may decrease the rewarding properties of AMPH. b, No differences in the frequency of cage crossing between pretests and posttests were noted in any group. *p < 0.05. Error bars indicate SEM.
AMPH-behanneling draacht DA frijlitting yn 'e NACC yn beide SN en PB-manlju
Der wiene gjin signifikante ferskillen tusken SN en PB-manlju yn 'e absolute bedraggen fan DA of DOPAC yn mikrodialysis baseline-samples (Fig. 2, ynsekten). AMPH-administraasje soarge foar signifikante ferheegingen yn extrazellulêre DA (F(12,108) = 8.42, p <0.001). De grutte en de doer fan dizze ferhegingen ferskilden lykwols net tusken SN- en PB-manlju - DA-nivo's wiene signifikant heger as basisline yn beide groepen foar elk fan 'e earste twa samplingperioden (40 min totaal) en kamen dan stadichoan werom nei baseline (Fig. 2, top). AMPH administration significantly decreased extracellular DOPAC in the NAcc in both SN and PB males (F(12,108) = 13.54, p < 0.001) and, again, these effects were similar in both groups. Neither SN nor PB males regained baseline levels before the end of sampling (Fig. 2, bottom).

- Download figuer
- Iepenje yn nije ljepper
- Download powerpoint
{kind=link}
Figure 2.
Levels of extracellular DA and DOPAC in the NAcc of SN and PB males following amphetamine treatment. Absolute amounts of DA and DOPAC in baseline dialysates did not differ between SN and PB males (insets). Peripheral amphetamine administration increased extracellular DA and decreased extracellular DOPAC within the NAcc. The magnitude of these responses was not different between SN and PB males. Arrows indicate times of injection. *Significant difference from the within-social experience baseline. Error bars indicate SEM.
AMPH-behanneling feroaret ferskillend D1R ferbinend yn 'e NACS fan SN en PB-manlju
Foarige ûndersiken hawwe bewize dat AMPH behannelings NAcc D1R gene en protein ekspresje ferbetterje (Liu et al., 2010). Additionally, pair-bonding experience elevates D1R binding (Aragona et al., 2006) yn 'e NAFC fan manlike pree-foles. Dêrom dogge wy hypotezearjen dat feroareingen yn DA-receptor ferbine yn 'e NACC kinne de gedrachsûntwikkeling tusken paadbonding en AMPH-belesting ûnderlizze. Wy ferwurke harsekrippen fan subjects brûkt yn de CPP-tests foar DA-receptor autoradiografyske binding. Twa-way-ANOVA-analyze hat oanjûn in wichtige ynteraksje tusken sosjale erfaring (SN of PB) en ynjeksje-type (saline of AMPH) op D1R-ferbining yn 'e NAcc (F(1,29) = 17.63, p <0.01). De post hoc test revealed that the densities of D1R binding in the NAcc of SN-AMPH and PB-saline groups were comparable and significantly higher than those of SN-saline and PB-AMPH groups (Fig. 3a). Neither AMPH treatment nor pair-bonding experience altered the density of D2R binding in the NAcc (Fig. 3b). In addition, no group differences were found in either D1R or D2R binding in the caudate–putamen (data are not shown).

- Download figuer
- Iepenje yn nije ljepper
- Download powerpoint
{kind=link}
Figure 3.
Pair bonding and AMPH interact to affect DA receptor binding. a, AMPH conditioning significantly increased the density of NAcc D1R binding in SN males (SN-AMPH) compared with saline-injected controls (SN-saline). However, PB males injected with saline (PB-saline) showed a level of D1R binding comparable to that of SN-AMPH males, and AMPH conditioning significantly decreased the density of NAcc D1R binding (PB-AMPH). b, No group differences were found in NAcc D2R binding. Photomicrographs illustrate D1R binding in the NAcc (circled) in the prairie vole brain. Bars with different Greek letters differ significantly from each other. Error bars indicate SEM. Scale bar, 1 mm.
NAcc D1R-aktivearring fertsjinnet AMPH-belje yn SN-manlju
Yn manlike prairiefollen, subkutane ynjeksjes fan in D1R, mar net D2R, antagonist foar AMPH-kondysje-sesjes eleminearre AMPH-induzearre CPP (Liu et al., 2010). Mei't de fereaske rol fan NAcc DA yn AMPH belje yn oare naget soarten (Yokel en Wise, 1975; Kehoe et al., 1996), we hypothesized that access to D1Rs in the NAcc during conditioning is essential for AMPH-induced CPP in SN male prairie voles. We found that SN males that received CSF injections into the NAcc displayed AMPH-induced CPP (t = 2.90, p <0.01) (Fig. 4). However, intra-NAcc administration of a D1R antagonist, SCH 23390, at either a low (4 ng/side) or high (100 ng/side) dose before conditioning sessions eliminated AMPH-induced CPP (Fig. 4). Yn tsjinstelling ta intra-NAcc-administraasje fan in D2R antagonist, etiklopride, op ien of in leech (4 ng / side; t = 3.25, p <0.01) as heech (100 ng / kant; t = 2.30, p < 0.05) dose did not block AMPH-induced CPP (Fig. 4). No differences were found in the frequencies of cage crosses between the pretests and posttests in any group, indicating no effect of treatment on locomotor activity (data are not shown).
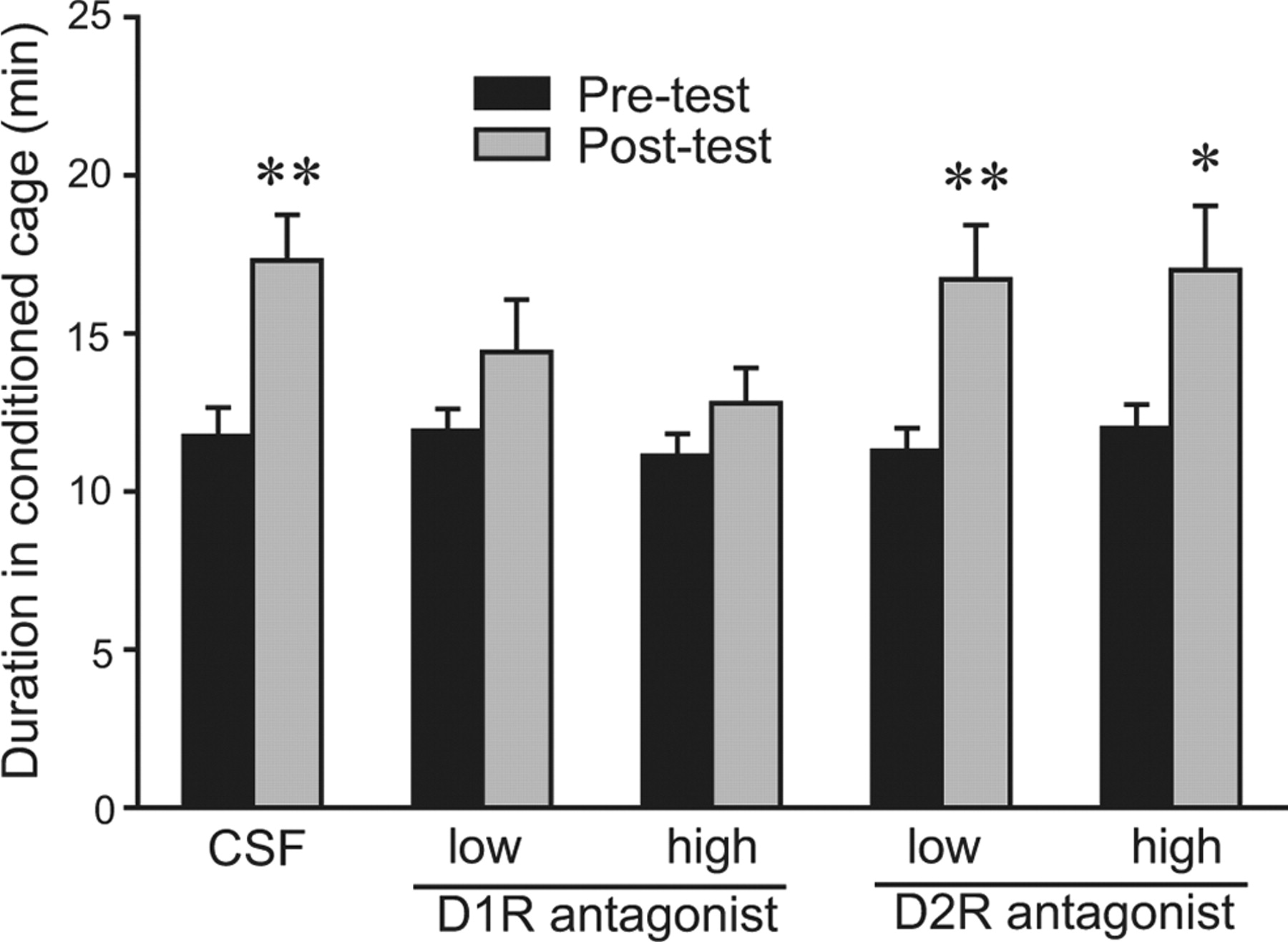
- Download figuer
- Iepenje yn nije ljepper
- Download powerpoint
{kind=link}
Figure 4.
The involvement of NAcc DA D1R and D2R in AMPH-induced CPP in sexually naive male prairie voles. All subjects received AMPH during conditioning sessions. On each of the 3 d of conditioning, 30 min before AMPH injections, subjects received bilateral intra-NAcc microinjections of either artificial CSF or CSF containing a low or high dose of SCH 23390 (D1R antagonist) or eticlopride (D2R antagonist). Subjects treated with CSF and either dose of the D2R antagonist formed an AMPH-induced CPP, i.e., they spent significantly more time in the conditioned cage during the posttest than during the pretest. However, treatment with either dose of the D1R antagonist eliminated AMPH-induced CPP, indicating that NAcc D1R, but not D2R, activation is essential for AMPH reinforcement in sexually naive male prairie voles. *p <0.05, **p < 0.01. Error bars indicate SEM.
Aktivaasje fan D1Rs yn 'e NAcc kin AMPH-induzearre CPP yn PB-manlju
Previous studies have shown that NAcc D1R activation is essential for AMPH-induced CPP and selective aggression, and that it impairs partner preference formation in male prairie voles (Aragona et al., 2006; Liu et al., 2010). Mei it each op de rol fan D1Rs yn dy gedrach en de fynst dat NAcc D1R ferbining is yn PB-AMPH-manlju dan yn PB-saline en SN-AMPH-manlju (Fig. 3a), we hypothesized that decreased D1R activity in the NAcc may be responsible for the lack of AMPH-induced CPP in PB males. To test this hypothesis, we injected CSF or CSF containing a D1R agonist or antagonist site-specifically into the NAcc before each of the three conditioning sessions and then tested for the presence of AMPH-induced CPP. As expected, PB males that received CSF injections did not show AMPH-induced CPP (Fig. 5). PB-manlju dy't lykwols intra-NAcc ynjeeksjes krigen fan 'e D1R agonist (t = 4.69, p <0.001), mar net antagonist, werjûn AMPH-feroarsake CPP (Fig. 5). There were no differences in the frequencies of cage crosses between the pretest and posttest for any group (data are not shown).

- Download figuer
- Iepenje yn nije ljepper
- Download powerpoint
{kind=link}
Figure 5.
Activation of the DA D1R in the NAcc enables AMPH-induced CPP in pair-bonded male prairie voles. All subjects were pair bonded and received AMPH during conditioning sessions. On each of the 3 d of conditioning, 30 min before AMPH injections, subjects received bilateral intra-NAcc injections of either CSF or CSF containing SFK 38393 (D1R agonist, 0.4 ng/side) or SCH 23390 (D1R antagonist, 4 ng/side). Subjects treated with CSF or a D1R antagonist did not form a CPP. However, activation of D1Rs with a D1R agonist induced CPP in pair-bonded male prairie voles. **p < 0.001. Error bars indicate SEM.
Diskusje
Studijen yn sawol minsk- en diermodellen hawwe in sterke relaasje tusken drugsbedriuw / misbrûk en sosjale gedrach (Jonge et al., 2011a). Because of its well known role in the generation of motivated behaviors, the mesolimbic DA system is in a key position to mediate the interaction between drugs of abuse and social behavior. We have recently demonstrated that repeated exposure to AMPH impairs pair-bond formation in male prairie voles and that NAcc DA mediates this effect (Liu et al., 2010). In the present study, we demonstrate that pair-bonding experience impairs AMPH-induced CPP and that this effect is also mediated by NAcc DA. Together, these studies demonstrate a reciprocal interaction between pair bonding and AMPH reward and suggest a role for NAcc DA in regulating such interactions.
Yn 'e aktuele stúdzje brûkten wy in earder fêststeld CPP-paradigm (Liu et al., 2010; Jonge et al., 2011b) to investigate the effects of pair-bonding experience on the rewarding properties of AMPH. We use the ambiguous term “rewarding properties” to describe the impact of AMPH on place conditioning because it allows us to simultaneously address the individual components of reward—including hedonics, associative learning, and incentive motivation (Berridge en Robinson, 2003) - dat binne yngien yn prosedure ûnderlizzende plakken kondysje (Hnasko et al., 2005; Wyt et al., 2005; Cunningham en Patel, 2007), without distinguishing between them. Our results demonstrate that AMPH conditioning induced a CPP in SN, but not PB, male voles, and as such, offer the first empirical evidence that pair-bonding experience decreases the rewarding properties of AMPH. Although this is the only study to investigate the effects of pair-bonding experience on the rewarding properties of drugs of abuse, previous studies have demonstrated that other social experiences/factors may also influence drug reward. For example, rats reared with multiple social cohorts self-administered less AMPH (Bardo et al., 2001) en kokaïne (Schenk et al., 1987) oer werhelle sesjes as raten allinich. Krekt lykwols waard de raten yn in feroverde omjouwing ûntstien dy't nije artikels en sosjale koördinaten befettet wat minder administraasjearbere AMPH, ferlernde self-administraasje geduld earder en ferplichte hegere dosinten fan AMPH om te reitsjen fan medikaminsjogger as paspoaten allinich (Schenk et al., 1988; Bardo et al., 1995, 2001; Green et al., 2002; Stappen et al., 2006). Fierder hawwe froulike raten oan 'e leverdruk foar kokaïne te folle minder responsen nei't se swier binne en folgjende partuering (Hecht et al., 1999), suggerearret dat it reproduktive proses en neurobiologyske wizigingen belutsen by it mêd fan 'e memmetaal kinne de fersterkjende eigenskippen fan kokaïne ôfnimme. Dizze begryp wurdt fierder stipe troch de befinings dat virginale rotten in foarkar hawwe foar in omjouwing dy't ferbûn binne mei kokaïne en maklik maklik útkomt mei in kokaïne-induzearre CPP (Seip et al., 2008), wylst lekkende dammen sterk foarkommen foar in omjouwing dy't ferbûn binne mei welpen oer ien assosjearre mei kokaïne (Mattson et al., 2001). Together, these studies imply that social factors may reduce the rewarding properties of psychostimulants. Our results extend these findings and demonstrate that pair-bonding experience impairs AMPH reward in prairie voles.
The rewarding properties of psychostimulant drugs of abuse, such as AMPH, are dependent on drug-induced increases in NAcc DA release and the subsequent activation of DA receptors (Yokel en Wise, 1975; Di Chiara en Imperato, 1988; Bergman et al., 1990; Kehoe et al., 1996). Therefore, alterations in either of these factors may underlie the effects of pair-bonding experience on AMPH reward. Levels of released DA have been closely correlated with the positive subjective effects of drugs of abuse (Volkow et al., 1999; Drevets et al., 2001; Leyton, 2010) and psychostimulant-induced NAcc DA release has been altered by social experience in some cases [e.g., isolation early in life (Kehoe et al., 1996; Kosten et al., 2005)], but not others [e.g., enriched social housing (Bardo et al., 1995)]. In the present study, AMPH treatment increased the level of extracellular DA in the NAcc, which is consistent with previous reports (Di Chiara en Imperato, 1988; Curtis en Wang, 2007; McKittrick en Abercrombie, 2007). Fierder feroarsake AMPH in direkte en langere ôfwiking op it nivo fan extrazellulêre DOPAC yn beide groepen. Dizze fynstelling is konsekwint mei dy fan eardere stúdzjes en de bekende rol fan AMPH yn 'e ynhibering fan monoaminoksidase - in enzyme dy't belutsen by de degradaasje fan DA (Groen en el Hait, 1978; Jones et al., 1998; Curtis en Wang, 2007). Yn beide gefallen wie it grutter en tydlike model fan ekstrazellulêre wizigingen yn 'e NACK fergelykber tusken SN en PB-manlju, wêrtroch't bepaald is dat it effekt is dat de effekten fan paadbondingen op AMPH-belje relatearre binne oan de frijlitting of metabolism fan NAcc DA.
Although pair-bonding experience did not influence AMPH-induced DA release or metabolism, it did influence the effects of AMPH on NAcc DA receptor binding (i.e., DA receptor expression and/or affinity). For example, consistent with previous studies, AMPH treatment increased NAcc D1R expression in SN males (Liu et al., 2010) and pair-bonding experience also elevated NAcc D1R binding (Aragona et al., 2006). However, the effect of AMPH on NAcc D1R binding was reversed in PB males compared with SN males, as PB males showed a significant decrease in D1R binding following AMPH treatment. No group differences were found in NAcc D2R binding in either the current or previous studies (Aragona et al., 2006; Liu et al., 2010). Together, these data demonstrate that pair-bonding experience and AMPH exposure each leads to D1R-specific changes within the NAcc of SN males. Further, as AMPH oppositely affected NAcc D1R binding in SN and PB voles, our data indicate that social bonding may be an important mediating factor in the effects of AMPH on the mesolimbic DA system. Correspondingly, cocaine has been found to induce a robust positive blood oxygenation level-dependent (BOLD) signal change throughout the mesolimbic DA system in virgin females, but a largely negative BOLD response in lactating dams, as measured by functional magnetic resonance imaging (Ferris et al., 2005), thus further indicating that social/sexual experience may play an important role in the neurobiological response to drugs of abuse. Alterations in receptor binding, such as those described above, may have profound effects on behavior because they modify the brain’s responsiveness to released neurotransmitters. In SN males, for example, AMPH-induced increases in D1R binding may play an important role during AMPH conditioning as intra-NAcc blockade of D1R, but not D2R, inhibited AMPH-induced CPP—a finding consistent with those in other species (Baker et al., 1998; Pitchers et al., 2010). Consequently, in PB males, decreased NAcc D1R binding may underlie the lack of an AMPH-induced CPP as NAcc D1R activation during AMPH conditioning enabled AMPH-induced CPP in PB males. Collectively, these data indicate that AMPH-induced decreases in NAcc D1R expression/affinity may underlie the effects of pair-bonding experience on AMPH reward.
Two interesting parallels between our findings and those from other studies that have investigated the relationship between drugs of abuse and social behavior are worth mentioning. First, pair bonding and repeated AMPH exposure each independently resulted in similar changes in the mesolimbic DA system of male prairie voles [i.e., enhanced D1R binding (Aragona et al., 2006) en ekspresje (Liu et al., 2010) in the NAcc]. Similarly, sexual experience and repeated exposure to psychostimulants each increased the density of dendritic spines on medium spiny neurons within the NAcc shell in rats (Robinson en Kolb, 1999; Pitchers et al., 2010). Fierder hat de pup oanwakke yn laitsjende damjen en kokaineadminte yn 'e jonkfroulju like ferlykbere patroanen fan positive BOLD-aktivearring yn it mesocorticolimbic systeem (Ferris et al., 2005). Together, these data support the notion that drugs of abuse usurp neural mechanisms and circuits that mediate adaptive behaviors (Panksepp et al., 2002). Second, our data suggest that the differential neurobiological responses to AMPH in SN and PB males may underlie group differences in AMPH-induced behavior. Similarly, lactating rats showed a suppression of activity within the mesocorticolimbic DA system in response to cocaine (as opposed to the increase in activity noted in virgin females) (Ferris et al., 2005) en in reduksje yn 'e kokaïne selsbehear (Hecht et al., 1999), fierder stipe fan 'e opmerking dat fysiologyske feroaringen dy't ferbân hâlde mei maatskiplik ûnderfining meie de leare eigenskippen fan misbrûk misbrûke.
De oanwêzigens fan sterke sosjale bondels yn 'e folwoeksenens kin de kwetsberens foar drugsmisbrûk ferminderje (Kosten et al., 1987). However, the neural mechanisms underlying this behavioral phenomenon are relatively unknown, perhaps due to the lack of an appropriate animal model. In the present study, we established the prairie vole as an animal model to investigate the neural mechanisms underlying the protective effects of adult social bonding on the vulnerability to drug abuse. Our findings indicate that pair-bonding experience decreases the rewarding properties of AMPH and that the mesolimbic DA system, particularly DA neurotransmission in the NAcc, mediates this effect. These findings, together with those from our previous study (Liu et al., 2010), in grûnslach foar takomstige ûndersyk opstelle fan de neurologyske meganisaasjes dy't de ferwidering fan relaasje tusken medisynferbân / misbrûk en sosjale bonding hawwe, dy't eventueel wichtige ynsjoch leverje kinne yn 'e previnsje of behanneling fan drugsmisbrûk.