
Komentarz: Wiązanie par sprawia, że używanie narkotyków jest mniej atrakcyjne dzięki mechanizmowi receptora dopaminy.
Abstrakcyjny
Chociaż ochronny wpływ więzi społecznych na używanie / nadużywanie narkotyków został dobrze udokumentowany, niewiele wiemy o podstawowych mechanizmach neuronalnych. Korzystanie z nornika preriowego (Microtus ochrogaster) — społecznie monogamiczny gryzoń, który tworzy długotrwałe więzi w parach po kojarzeniu — pokazujemy, że warunkowanie amfetaminą (AMPH) wywołało warunkową preferencję miejsca (CPP) u naiwnych płciowo (SN), ale nie związanych parą (PB), samców. Chociaż leczenie AMPH indukowało podobną wielkość uwalniania dopaminy w jądrze półleżącym (NAcc) samców SN i PB, miało ono zróżnicowany wpływ na wiązanie receptora NAcc D1 (D1R). Konkretnie, leczenie AMPH zwiększyło wiązanie D1R u SN, ale zmniejszyło wiązanie D1R u samców PB. Antagonizm NAcc D1R, ale nie receptora D2, blokował CPP indukowany przez AMPH u samców SN, a aktywacja NAcc D1R przed kondycjonowaniem AMPH umożliwiła CPP indukowaną przez AMPH u samców PB. Razem nasze dane pokazują, że doświadczenie wiązania par zmniejsza satysfakcjonujące właściwości AMPH dzięki mechanizmowi, w którym pośredniczy D1R.
Wprowadzenie
Powszechnie wiadomo, że zażywanie narkotyków i nadużywanie mają poważne konsekwencje dla różnorodnych zachowań społecznych, w tym więzi społecznych (Young i in., 2011a). Odwrotnie, czynniki społeczne, takie jak obecność lub brak silnych więzi społecznych w okresie rozwoju lub w okresie dorosłości, mogą wpływać na używanie narkotyków i podatność na nadużywanie narkotyków. Na przykład silne przywiązania między młodzieżą a rodzicami wiązały się ze zmniejszonymi prawdopodobieństwem problemów związanych z używaniem substancji (Ellickson i in., 1999; Bell i wsp., 2000). Ponadto u osób, które są już uzależnione, bliskie relacje między małżonkami pomagają w wyzdrowieniu z narkomanii (Kosten i in., 1987). W przeciwieństwie do tego słabe przywiązania społeczne mogą zwiększać podatność na nadużywanie substancji, ponieważ niepewne style przywiązania dorosłych są pozytywnie związane z alkoholem i nielegalnym nadużywaniem substancji (Brennan i Shaver, 1995; Vungkhanching i in., 2004; Caspers i in., 2005).
Mechanizmy neuronalne leżące u podstaw interakcji między więziami społecznymi a używaniem/nadużywaniem narkotyków są nadal w dużej mierze nieznane; jednak może być zaangażowany układ mezolimbiczny dopaminy (DA), w szczególności neuroprzekaźnictwo DA w jądrze półleżącym (NAcc) (Young i in., 2011a). Badania w nornicy preriowej (Microtus ochrogaster) - społecznie monogamiczny gryzoń, który tworzy trwałe więzi między dorosłymi partnerami (tj. Więzi par) (Insel and Hulihan, 1995; Mattson i in., 2001) - pokazali, że NAcc DA pośredniczy zarówno w tworzeniu, jak i utrzymywaniu wiązań parowych (Gingrich i in., 2000; Aragona i in., 2003, 2006). Dodatkowo neurotransmisja NAcc DA jest aktywowana przez wszystkie znane leki nadużywające (Di Chiara i Imperato, 1988; Bergman i in., 1990; Koob i Nestler, 1997; Nestler, 2005) i powtarzająca się ekspozycja na leki psychostymulujące zmienia uwalnianie DA i wrażliwość receptora, jak również morfologię neuronów NAcc (Henry i in., 1989; Robinson i Kolb, 1999). Uważa się, że te zmiany leżą u podstaw modyfikacji zachowań wywołanych przez narkotyki (Robinson i Becker, 1986), w tym zachowania społeczne (Fiorino i Phillips, 1999).
Ponieważ zarówno zachowanie wiązania par, jak i jego regulacja przez neuroprzekaźnictwo DA w NAcc zostały dobrze scharakteryzowane u nornika preriowego (Gingrich i in., 2000; Aragona i in., 2003, 2006), podjęto niedawne wysiłki, aby uczynić ten gatunek modelem zwierzęcym, w którym można zbadać interakcje behawioralne między więziami społecznymi a używaniem/nadużywaniem narkotyków, a także leżące u ich podstaw mechanizmy neuronalne. Wykazano, że amfetamina (AMPH) ma właściwości nagradzające u norników preriowych, ponieważ kondycjonowanie AMPH wywołuje warunkową preferencję miejsca (CPP) u obu płci (Aragona i in., 2007; Liu i wsp., 2010; Young i in., 2011b); w procesie tym pośredniczy neurotransmisja DA w NAcc (Curtis i Wang, 2007; Liu i wsp., 2010). Co najciekawsze, powtarzana ekspozycja na AMPH hamuje powstawanie wywołanych kojarzeniem preferencji partnerów u samców nornicy preriowej, a to wywołane przez AMPH upośledzenie wiązania par jest regulowane, przynajmniej częściowo, przez aktywację receptora NAcc DA typu D1 (D1R) (Liu i wsp., 2010). W niniejszym badaniu zbadaliśmy wpływ doświadczenia wiązania par na CPP wywołane przez AMPH. Postawiliśmy hipotezę, że doświadczenie wiązania par zmniejszy nagradzające efekty AMPH i że NAcc DA może być zaangażowany w to zjawisko behawioralne.
Materiały i Metody
PRZEDMIOTÓW.
Przedmioty były męskimi norami preriowymi (M. ochrogaster) z laboratoryjnej kolonii hodowlanej. Badani zostali odsadzeni w wieku 21 dni i trzymani w parach rodzeństwa tej samej płci w plastikowych klatkach (12 × 28 × 16 cm). Zapewniono wodę i jedzenie ad libitum. Wszystkie klatki utrzymywano w cyklu światło:ciemność 14:10, a temperaturę utrzymywano na poziomie 20°C. Badani w wieku około 75 dni byli albo stale trzymani ze swoim rodzeństwem tej samej płci [a tym samym utrzymywani naiwnie seksualnie (SN)] lub sparowani z niespokrewnioną nienaruszoną samicą przez 2 tygodnie, aby związać się w parę (PB). Zarówno osoby z SN, jak i PB były badane w wieku około 90 dni.
KONDYCJONOWANIE AMPH I TESTOWANIE CPP.
Procedury te przeprowadzono zgodnie z wcześniejszym opisem (Liu i wsp., 2010; Young i in., 2011b). Krótko mówiąc, aparat do badania CPP składał się z dwóch klatek (12 × 28 × 16 cm), jednej czarnej z metalowym wierzchem i jednej białej z wierzchem z siatki, połączonych pustą rurką (7.5 × 16 cm). Chociaż norniki preriowe na ogół wolą biel niż ciemną klatkę (Aragona i in., 2007), istnieje wiele indywidualnych różnic w tej preferencji. Dlatego pierwszego dnia przetestowaliśmy wszystkie osoby pod kątem ich początkowych preferencji klatkowych podczas 1-minutowego testu wstępnego. Podczas tego testu wszyscy badani mieli swobodny dostęp do obu klatek i obliczyliśmy ilość czasu, jaki każdy osobnik spędził w każdej klatce. W dniach 30–2 badani otrzymywali dwie 4-minutowe sesje kondycjonujące w odstępie 40 godzin. W sesji porannej badani otrzymali albo 6 mg/kg AMPH (Sigma) rozpuszczony w 1.0% soli fizjologicznej (grupy SN-AMPH i PB-AMPH) albo sam roztwór soli (grupy SN-sól fizjologiczna i PB-sól fizjologiczna) i zostali umieszczeni w klatce w którym spędzili mniej czasu podczas testu wstępnego (klatka kondycjonowana). W sesji popołudniowej wszyscy badani otrzymali zastrzyk soli fizjologicznej i zostali umieszczeni w drugiej klatce. W dniu 0.9, osobniki były ponownie testowane pod kątem preferencji w klatkach w 5-minutowym teście post-testowym. Bezpośrednio po teście post-teście, osobnikom szybko ścięto głowy, a ich mózgi zamrożono na suchym lodzie. Skrawki mózgu poddano następnie obróbce pod kątem autoradiograficznego wiązania receptora typu D30R i DA D1 (D2R).
MIKRODIALIZA MÓZGU I ANALIZA WYKRYWANIA ELEKTROCHEMICZNEGO HPLC.
Sondy do mikrodializy skonstruowano w sposób opisany powyżej (Curtis i Wang, 2007) i wszczepiono do lewego NAcc (współrzędne stereotaktyczne z bregma: przednia 2.1 mm, boczna 0.6 mm, brzuszna 6.3 mm) w znieczuleniu pentabarbitolem sodu (1 mg/10 kg masy ciała). Zwierzęta pozostawiono do regeneracji przez noc, a następnego ranka poddano je testom. Sondy perfundowano w sposób ciągły przy 2.3 μl/min roztworem izotonicznym dla sodu (144 mm NaCl), potas (2.8 m³mKCl), wapń (1.2 m³m CaCl2) i magnez (0.9 mm MgCl2) (Sved i Curtis, 1993).
Po całonocnym odzyskiwaniu, cztery 20-minutowe próbki linii podstawowej zebrano do fiolek zawierających 5 μl 0.1N kwasu nadchlorowego. Następnie osobnicy otrzymywali dootrzewnowe wstrzyknięcie AMPH (1.0 mg/kg) i próbki dializatu pobierano w sposób ciągły w 20-minutowych odstępach przez 3 godziny. Próbki dializatu natychmiast zamrożono w -80°C do czasu analizy. Ilości DA i DOPAC w każdej próbce określono metodą HPLC z detekcją elektrochemiczną (ECD), jak opisano wcześniej (Curtis i Wang, 2007). Pod koniec okresu pobierania próbek osobniki zostały zabite, aby ocenić umieszczenie sondy.
AUTORADIOGRAFIA RECEPTORA DA.
Skrawki mózgu (20 μm) w odstępach 120 μm poddano obróbce pod kątem autoradiograficznego wiązania receptora DA przy użyciu ustalonej metody (Aragona i in., 2006). W skrócie, sekcje zostały wypłukane w 50 mm Tris-HCl, pH 7.4 i inkubowany w 50 mm Bufor jonowy Tris-HCl zawierający 120 mm NaCl, 5 mm KCl, 2 mm CaCl2i 1 mlnm MgCl2 z albo [125I]SCH 23982 (ligand D1R) lub [125I]2'-jodospiperon (ligand D2R) (PerkinElmer). Następnie skrawki utrwalono w 0.1% paraformaldehydzie i dokładnie przepłukano w buforze jonowym Tris-HCl. Slajdy zanurzono w wodzie destylowanej, osuszono i wystawiono na kliszę BioMax MR (Kodak) w celu wygenerowania autoradiogramów. Gęstości optyczne wiązania D1R i D2R w NAcc i skorupie ogoniastej (CP) określono ilościowo w trzech dopasowanych anatomicznie wycinkach mózgu na zwierzę na podstawie autoradiogramów przy użyciu skomputeryzowanego programu do obrazowania (NIH Image 1.64).
KANULACJA I MIKROWSTRZYKIWANIE STEREOTAKSYCZNE.
Badani byli znieczulani pentobarbitalem sodu, a 26-gauge dwustronne kaniule ze stali nierdzewnej (Plastic One) wszczepiono stereotaktycznie i wycelowano w NAcc, jak opisano wcześniej (Aragona i in., 2003). Badani mogli wyzdrowieć przez 3–7 dni. W każdym z 3 dni kondycjonowania, 30 minut przed wstrzyknięciami AMPH, badani otrzymywali mikrowstrzyknięcia sztucznego płynu mózgowo-rdzeniowego (200 ml/stronę) lub płynu mózgowo-rdzeniowego zawierającego agonistę D1R SKF 38393, antagonistę D1R SCH 23390 lub antagonistę D2R etyklopryd (Sigma ). Po teście CPP wszystkim badanym szybko odcięto głowy, a ich mózgi pobrano w celu histologicznej weryfikacji miejsc wstrzyknięć. Osoby z niewłaściwie umieszczonymi kaniulami zostały wykluczone z analizy danych.
KWANTYFIKACJA DANYCH I ANALIZA STATYSTYCZNA.
CPP określono za pomocą sparowanych próbek t test porównujący ilość czasu, jaką badani spędzili w klimatyzowanej klatce między testami wstępnymi i testami końcowymi. Wpisy w klatkach między pretestami i posttestami zostały również przeanalizowane przez t test, aby ocenić, czy AMPH, agonista D1R, D1R lub antagonista D2R wpływa na aktywność lokomotoryczną. Bezwzględne ilości wyjściowej DA i DOPAC w dializatach porównano między grupami za pomocą t test. W celu oceny wpływu AMPH w czasie, ilości DA i DOPAC w każdej próbce wyjściowej i po AMPH wyrażono jako procent średniej ilości wyjściowej. Wartości te zostały następnie przeanalizowane za pomocą powtarzanych pomiarów ANOVA, a następnie Student-Neuman-Keuls (SNK) post hoc test. Na koniec, różnice grupowe w gęstościach wiązania D1R i D2R w NAcc i CP analizowano za pomocą dwukierunkowej ANOVA, a następnie SNK post hoc test.
PROJEKT EKSPERYMENTALNY.
Eksperyment 1 został zaprojektowany, aby ujawnić wpływ doświadczenia wiązania par na CPP wywołane przez AMPH. Samce SN i PB zostały wstępnie przebadane w aparacie CPP. Następnie podzielono ich na cztery grupy, które otrzymały zastrzyki z soli fizjologicznej (n = 5 dla samców SN i n = 9 dla samców PB) lub AMPH (1.0 mg/kg; n = 8 dla samców SN i n = 8 dla samców PB) podczas porannych sesji kondycjonowania przez następne 3 dni (Liu i wsp., 2010). Następnie wszyscy badani otrzymali posttest CPP.
Eksperyment 2 porównał uwalnianie DA indukowane przez AMPH w NAcc między SN (n = 6) i PB (n = 5) mężczyźni. Osobnikom wszczepiono sondę do mikrodializy wycelowaną w NAcc. Po całonocnej regeneracji z ciągłą perfuzją izotonicznego roztworu przez sondy, zebrano cztery próbki dializatu o długości 20 min. Następnie badani otrzymywali dootrzewnową iniekcję AMPH (1.0 mg/kg) i próbki dializatu pobierano w sposób ciągły co 20 minut przez 3 godziny. Próbki te były następnie analizowane pod kątem stężeń DA i DOPAC za pomocą analizy HPLC-ECD (Curtis i Wang, 2007).
W eksperymencie 3 zbadano wpływ interakcji między wiązaniem par i traktowaniem AMPH na wiązanie receptora DA w NAcc. Skrawki mózgu osobników badanych w eksperymencie 1 poddano obróbce pod kątem wiązania D1R i D2R przy użyciu autoradiografii receptora.
W eksperymencie 4 zbadano rolę receptorów NAcc DA w CPP indukowanym przez AMPH. Samcom SN wszczepiono kaniule prowadzące obustronnie skierowane w NAcc. Po 3 dniach powrotu do zdrowia, badani otrzymali wstępny test CPP, a następnie zostali losowo przypisani do jednej z pięciu grup eksperymentalnych [wstrzyknięcia płynu mózgowo-rdzeniowego do NAcc (n = 8) lub do-NAcc wstrzyknięcia płynu mózgowo-rdzeniowego zawierającego niski (4 ng/stronę); n= 8) lub wysoki (100 ng/stronę); n = 6) dawka antagonisty D1R, SCH 23390, lub niska (4 ng/stronę); n = 8) lub wysoki (100 ng/stronę); n = 7) dawka antagonisty D2R, etykloprydu]. Trzydzieści minut później badani otrzymali zastrzyk AMPH (1.0 mg/kg, ip). Procedurę tę powtarzano przez 3 kolejne dni podczas kondycjonowania AMPH. Następnie badani otrzymali posttest CPP.
W eksperymencie 5 zbadano rolę NAcc D1R w pośredniczeniu w CPP indukowanym przez AMPH u samców PB. Osoby z PB podzielono na trzy grupy, otrzymujące zastrzyki z płynu mózgowo-rdzeniowego do NAcc (n = 10) lub CSF zawierający agonistę D1R, SKF 38393 (0.4 ng/stronę; n = 12) lub antagonista D1R, SCH 23390 (4 ng/stronę; n = 10), przed kondycjonowaniem AMPH. Kaniulacja mózgu, zastrzyki AMPH i testy CPP były takie same, jak opisano w eksperymencie 4.
Efekt
Doświadczenie wiązania par zmniejsza satysfakcjonujące właściwości AMPH
W naszym poprzednim badaniu leczenie AMPH zaburzało preferencje partnerów wywołane kojarzeniem się samców norników preriowych, co wskazuje na hamujący wpływ ekspozycji na AMPH na zachowanie związane z wiązaniem się par (Liu i wsp., 2010). W niniejszym badaniu przetestowaliśmy wzajemną zależność: efekty doświadczenia łączenia par na nagrodę AMPH. Trzy dni kondycjonowania 1.0 mg / kg AMPH wywołały CPP u mężczyzn SN (t = 2.45, p <0.05), ale nie u samców, którzy byli sparowani z kobietą przez 2 tygodnie (tj. Samce PB) (Rys. 1a). Zastrzyki z soli fizjologicznej nie miały wpływu w żadnej z grup. Co ważne, nie znaleziono różnic w częstotliwościach przekraczania klatek przez zwierzęta między testami wstępnymi i po testach, co sugeruje, że upośledzenie CPP u samców PB nie było spowodowane zmienioną aktywnością lokomotoryczną podczas testu behawioralnego (Rys. 1b).

- Pobierz rysunek
- Otwórz w nowej karcie
- Pobierz Powerpoint
{kind=link}
Rysunek 1.
Kondycjonowanie AMPH indukuje CPP u samców norników preriowych SN, ale nie PB. a, samce SN lub PB, które otrzymały sól fizjologiczną (odpowiednio sól fizjologiczną SN lub sól fizjologiczną PB) podczas 3 dni kondycjonowania nie wytworzyły CPP, tj. spędziły równe ilości czasu w kondycjonowanej klatce podczas 30-minutowych testów wstępnych i po testach. Kondycjonowanie AMPH wywołało CPP u samców SN (SN-AMPH), ale nie u samców PB (PB-AMPH), wskazując, że doświadczenie wiązania par może zmniejszyć nagradzające właściwości AMPH. b, W żadnej grupie nie stwierdzono różnic w częstotliwości krzyżowania klatek między testami wstępnymi i potestowymi. *p < 0.05. Słupki błędów wskazują SEM.
Leczenie AMPH indukuje uwalnianie DA w NAcc zarówno u mężczyzn SN, jak i PB
Nie stwierdzono istotnych różnic między mężczyznami SN i PB w bezwzględnych ilościach DA lub DOPAC w próbkach podstawowych mikrodializy (Rys. 2, wstawki). Podawanie AMPH powodowało znaczący wzrost pozakomórkowej DA (F(12,108) = 8.42, p <0.001). Jednak wielkość i czas trwania tych wzrostów nie różniły się między samcami SN i PB - poziomy DA były znacznie wyższe niż wartość wyjściowa w obu grupach dla każdego z pierwszych dwóch okresów pobierania próbek (łącznie 40 minut), a następnie powoli powracały do wartości wyjściowej (Rys. 2, szczyt). Podawanie AMPH znacząco zmniejszyło zewnątrzkomórkowy DOPAC w NAcc zarówno u samców SN, jak i PB (F(12,108) = 13.54, p < 0.001) i ponownie efekty te były podobne w obu grupach. Ani samce SN, ani PB nie odzyskały poziomu wyjściowego przed końcem pobierania próbek (Rys. 2, na dole).

- Pobierz rysunek
- Otwórz w nowej karcie
- Pobierz Powerpoint
{kind=link}
Rysunek 2.
Poziomy zewnątrzkomórkowej DA i DOPAC w NAcc samców SN i PB po leczeniu amfetaminą. Bezwzględne ilości DA i DOPAC w wyjściowych dializatach nie różniły się między samcami SN i PB (wstawki). Obwodowe podawanie amfetaminy zwiększyło zewnątrzkomórkową DA i zmniejszyło zewnątrzkomórkowy DOPAC w obrębie NAcc. Wielkość tych odpowiedzi nie różniła się między samcami SN i PB. Strzałki wskazują czasy iniekcji. *Istotna różnica w stosunku do poziomu wyjściowego doświadczeń w obrębie społeczności. Słupki błędów wskazują SEM.
Leczenie AMPH w różny sposób zmienia wiązanie D1R w NAcc samców SN i PB
Poprzednie badania wykazały, że leczenie AMPH wzmacnia ekspresję genu i białka NAcc D1R (Liu i wsp., 2010). Dodatkowo doświadczenie wiązania par podnosi jakość wiązania D1R (Aragona i in., 2006) w NAcc męskich norników preriowych. Dlatego postawiliśmy hipotezę, że zmiany w wiązaniu receptora DA w NAcc mogą leżeć u podstaw interakcji behawioralnej między wiązaniem par a nagrodą AMPH. Przetworzyliśmy skrawki mózgu od osób wykorzystanych w testach CPP pod kątem wiązania autoradiograficznego receptora DA. Dwuczynnikowa analiza ANOVA wykazała znaczącą interakcję między doświadczeniem społecznym (SN lub PB) a typem zastrzyku (solanka lub AMPH) na wiązaniu D1R w NAcc (F(1,29) = 17.63, p <0.01). Plik post hoc test wykazał, że gęstości wiązania D1R w NAcc grup SN-AMPH i PB-solanka były porównywalne i znacznie wyższe niż w grupach SN-solanka i PB-AMPH (Rys. 3a). Ani traktowanie AMPH, ani doświadczenie wiązania par nie zmieniły gęstości wiązania D2R w NAcc (Rys. 3b). Ponadto nie stwierdzono różnic między grupami w wiązaniu D1R lub D2R w jądrze ogoniastym-powłoka (dane nie są pokazane).

- Pobierz rysunek
- Otwórz w nowej karcie
- Pobierz Powerpoint
{kind=link}
Rysunek 3.
Wiązanie par i AMPH oddziałują, wpływając na wiązanie receptora DA. a, kondycjonowanie AMPH znacząco zwiększyło gęstość wiązania NAcc D1R u samców SN (SN-AMPH) w porównaniu z kontrolami, którym wstrzyknięto sól fizjologiczną (SN-solanka). Jednak samce PB, którym wstrzyknięto sól fizjologiczną (sól fizjologiczna PB) wykazywały poziom wiązania D1R porównywalny do tego u samców SN-AMPH, a kondycjonowanie AMPH znacząco zmniejszyło gęstość wiązania NAcc D1R (PB-AMPH). b, Nie stwierdzono różnic między grupami w wiązaniu NAcc D2R. Fotomikrografie ilustrują wiązanie D1R w NAcc (zakreślone) w mózgu nornika preriowego. Słupki z różnymi literami greckimi znacznie różnią się od siebie. Słupki błędów wskazują SEM. Skala, 1 mm.
Aktywacja NAcc D1R pośredniczy w nagradzaniu AMPH u mężczyzn SN
W męskich nornicach preriowych wstrzyknięcia podskórne D1R, ale nie antagonisty D2R, przed sesjami kondycjonowania AMPH wyeliminowały CPP indukowane przez AMPH (Liu i wsp., 2010). Biorąc pod uwagę ustaloną rolę DA NAcc w nagrodzie AMPH u innych gatunków gryzoni (Yokel and Wise, 1975; Kehoe i in., 1996), postawiliśmy hipotezę, że dostęp do D1R w NAcc podczas kondycjonowania jest niezbędny dla wywołanego przez AMPH CPP u samców norników preriowych SN. Odkryliśmy, że samce SN, które otrzymały wstrzyknięcia płynu mózgowo-rdzeniowego do NAcc, wykazywały CPP wywołane przez AMPH (t = 2.90, p <0.01) (Rys. 4). Jednakże, podanie wewnątrz NAcc antagonisty D1R, SCH 23390, w niskiej (4 ng/stronę) lub wysokiej (100 ng/stronę) dawce przed sesjami kondycjonowania eliminowało CPP wywoływane przez AMPH (Rys. 4). W przeciwieństwie do tego, podawanie wewnątrz-NAcc antagonisty D2R, etliklopidu, albo w niskim (4 ng / bok; t = 3.25, p <0.01) lub wysoki (100 ng / bok; t = 2.30, p < 0.05) dawka nie blokowała CPP indukowanej przez AMPH (Rys. 4). Nie stwierdzono różnic w częstości krzyżowania klatek między testami wstępnymi i testami końcowymi w żadnej grupie, co wskazuje na brak wpływu leczenia na aktywność lokomotoryczną (dane nie są pokazane).
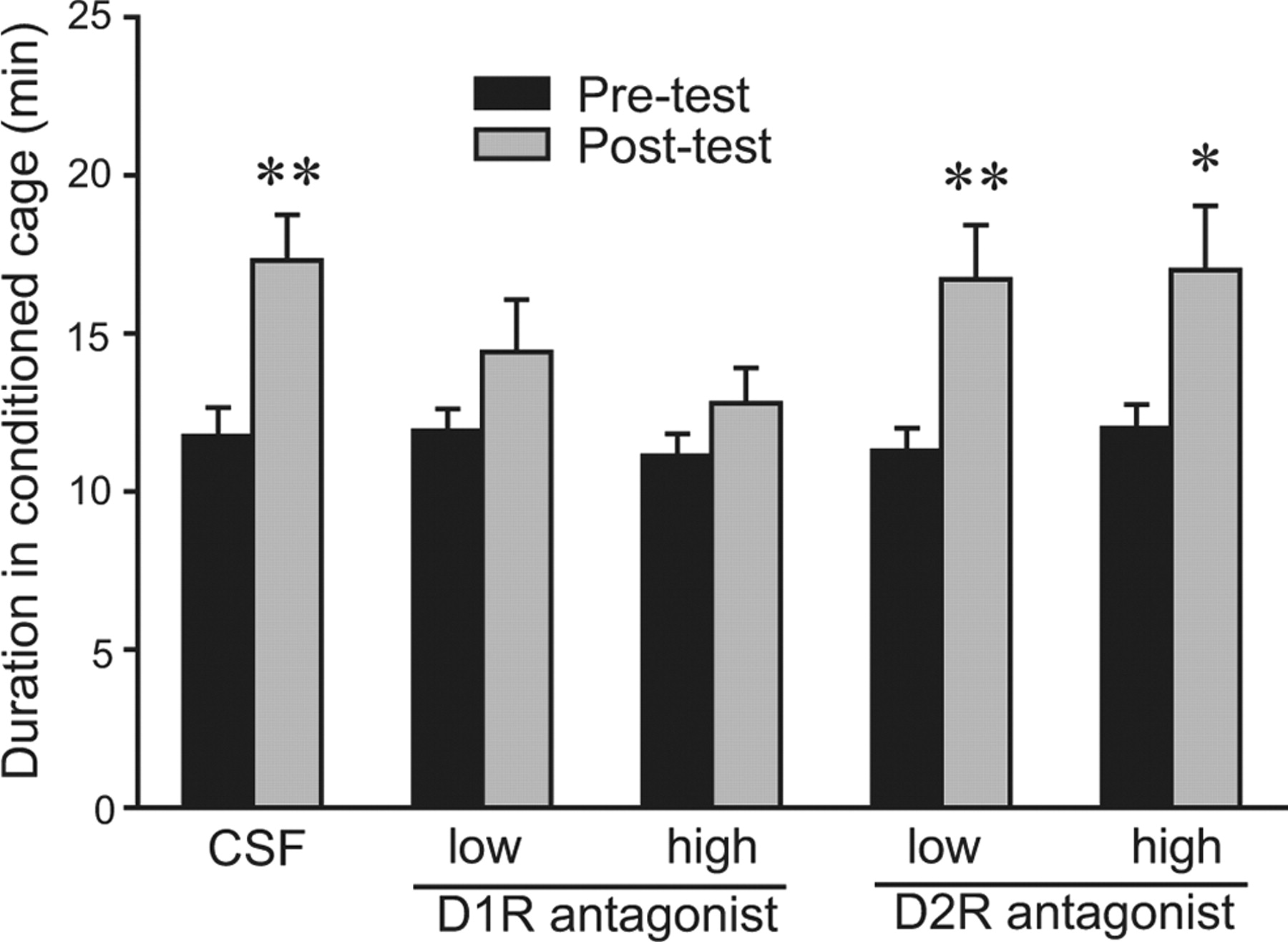
- Pobierz rysunek
- Otwórz w nowej karcie
- Pobierz Powerpoint
{kind=link}
Rysunek 4.
Zaangażowanie NAcc DA D1R i D2R w CPP indukowanym przez AMPH u naiwnych płciowo samców norników preriowych. Wszyscy badani otrzymywali AMPH podczas sesji kondycyjnych. W każdym z 3 dni kondycjonowania, 30 minut przed wstrzyknięciami AMPH, osobnicy otrzymywali obustronne mikroiniekcje do-NAcc sztucznego płynu mózgowo-rdzeniowego lub płynu mózgowo-rdzeniowego zawierającego niską lub wysoką dawkę SCH 23390 (antagonista D1R) lub etyklopryd (antagonista D2R). Osobnicy leczeni CSF i dowolną dawką antagonisty D2R wytworzyli CPP indukowaną przez AMPH, tj. spędzili znacznie więcej czasu w kondycjonowanej klatce podczas testu post-test niż podczas testu wstępnego. Jednak leczenie dowolną dawką antagonisty D1R eliminowało CPP indukowane przez AMPH, co wskazuje, że aktywacja NAcc D1R, ale nie D2R, jest niezbędna do wzmocnienia AMPH u naiwnych płciowo samców nornicy preriowej. *p <0.05, **p < 0.01. Słupki błędów wskazują SEM.
Aktywacja D1R w NAcc umożliwia CPP indukowane przez AMPH u mężczyzn PB
Wcześniejsze badania wykazały, że aktywacja NAcc D1R jest niezbędna dla wywołanej przez AMPH CPP i selektywnej agresji oraz że upośledza tworzenie preferencji partnerów u samców norników preriowych (Aragona i in., 2006; Liu i wsp., 2010). Biorąc pod uwagę rolę D1R w tych zachowaniach i odkrycie, że wiązanie NAcc D1R jest niższe u samców PB-AMPH niż u samców PB-solanki i SN-AMPH (Rys. 3a), postawiliśmy hipotezę, że zmniejszona aktywność D1R w NAcc może być odpowiedzialna za brak CPP indukowanego przez AMPH u samców PB. Aby przetestować tę hipotezę, wstrzyknęliśmy CSF lub CSF zawierające miejscowo agonistę lub antagonistę D1R do NAcc przed każdą z trzech sesji kondycjonowania, a następnie testowaliśmy na obecność CPP indukowanego przez AMPH. Zgodnie z oczekiwaniami, samce PB, które otrzymały zastrzyki z płynu mózgowo-rdzeniowego, nie wykazywały CPP wywołanego przez AMPH (Rys. 5). Jednak mężczyźni PB, którzy otrzymywali wstrzyknięcia wewnątrz-NAcc agonisty D1R (t = 4.69, p <0.001), ale nie antagonista, wykazywał CPP indukowany przez AMPH (Rys. 5). Nie było różnic w częstości krzyżowania klatek między testem wstępnym i posttestowym dla żadnej grupy (dane nie są pokazane).

- Pobierz rysunek
- Otwórz w nowej karcie
- Pobierz Powerpoint
{kind=link}
Rysunek 5.
Aktywacja DA D1R w NAcc umożliwia wywołane przez AMPH CPP u połączonych parami samców norników preriowych. Wszyscy badani byli związani parami i otrzymywali AMPH podczas sesji kondycjonujących. W każdym z 3 dni kondycjonowania, 30 minut przed wstrzyknięciami AMPH, pacjenci otrzymywali obustronne wstrzyknięcia do NAcc CSF lub CSF zawierające SFK 38393 (agonista D1R, 0.4 ng/stronę) lub SCH 23390 (antagonista D1R, 4 ng/stronę ). Osobnicy leczeni CSF lub antagonistą D1R nie tworzyli CPP. Jednak aktywacja D1R za pomocą agonisty D1R wywołała CPP u połączonych parami samców norników preriowych. **p < 0.001. Słupki błędów wskazują SEM.
Dyskusja
Badania na ludziach i modelach zwierzęcych wykazały silny związek między zażywaniem / nadużywaniem narkotyków a zachowaniem społecznym (Young i in., 2011a). Ze względu na swoją dobrze znaną rolę w generowaniu motywowanych zachowań, mezolimbiczny system DA ma kluczowe znaczenie dla pośredniczenia w interakcji między nadużywanymi narkotykami a zachowaniami społecznymi. Niedawno wykazaliśmy, że wielokrotna ekspozycja na AMPH osłabia tworzenie wiązań par u samców norników preriowych i że NAcc DA pośredniczy w tym efekcie (Liu i wsp., 2010). W niniejszym badaniu pokazujemy, że doświadczenie wiązania par osłabia CPP indukowane przez AMPH i że w tym efekcie pośredniczy również NAcc DA. Razem, badania te wykazują wzajemną interakcję między wiązaniem par a nagrodą AMPH i sugerują rolę NAcc DA w regulowaniu takich interakcji.
W bieżącym badaniu zastosowaliśmy wcześniej ustalony paradygmat CPP (Liu i wsp., 2010; Young i in., 2011b) w celu zbadania wpływu doświadczenia wiązania par na nagradzające właściwości AMPH. Używamy niejednoznacznego terminu „właściwości nagradzania”, aby opisać wpływ AMPH na warunkowanie miejsca, ponieważ pozwala nam to jednocześnie zająć się poszczególnymi składnikami nagrody – w tym hedoniką, uczeniem się asocjacyjnym i motywacją motywacyjną (Berridge i Robinson, 2003) - które były związane z procesami warunkującymi miejsce (Hnasko i in., 2005; White i in., 2005; Cunningham i Patel, 2007), bez rozróżniania między nimi. Nasze wyniki pokazują, że kondycjonowanie AMPH indukowało CPP w SN, ale nie PB, samce norników, i jako takie dostarczają pierwszego empirycznego dowodu na to, że doświadczenie wiązania par zmniejsza nagradzające właściwości AMPH. Chociaż jest to jedyne badanie, w którym zbadano wpływ doświadczenia związanego z łączeniem się w pary na nagradzające właściwości nadużywanych narkotyków, poprzednie badania wykazały, że inne doświadczenia/czynniki społeczne mogą również wpływać na nagrodę za narkotyki. Na przykład szczury hodowane z wieloma kohortami społecznymi samodzielnie podawały mniej AMPH (Bardo i in., 2001) i kokaina (Schenk i in., 1987) przez powtarzające się sesje niż szczury hodowane same. Podobnie, szczury hodowane w wzbogaconym środowisku, które zawierały nowe przedmioty i kohorty społeczne, które samodzielnie podawały mniej AMPH, wcześniej wygasły zachowania związane z samopodawaniem i wymagały wyższych dawek AMPH w celu przywrócenia poszukiwania narkotyków niż szczury trzymane samotnie (Schenk i in., 1988; Bardo i in., 1995, 2001; Green i wsp., 2002; Stairs i in., 2006). Ponadto samice szczurów przeszkolonych w zakresie nacisku na kokainę wykazały znacznie mniej odpowiedzi po zajściu w ciążę i po porodzie (Hecht i in., 1999), sugerując, że proces reprodukcyjny i zmiany neurobiologiczne związane z doświadczeniem matki mogą zmniejszyć wzmacniające właściwości kokainy. Pogląd ten jest dodatkowo poparty odkryciem, że pierwotne szczury preferowały środowisko związane z kokainą i łatwo wyrażały CPP wywołane kokainą (Seip i in., 2008), podczas gdy matki w okresie laktacji zdecydowanie preferowały środowisko związane ze szczeniętami w porównaniu z kokainą (Mattson i in., 2001). Razem, badania te sugerują, że czynniki społeczne mogą zmniejszać nagradzające właściwości psychostymulantów. Nasze wyniki poszerzają te odkrycia i pokazują, że doświadczenie wiązania par osłabia nagrodę AMPH u norników preriowych.
Nagradzające właściwości nadużywanych leków psychostymulujących, takich jak AMPH, zależą od wywołanego lekiem wzrostu uwalniania DA NAcc i późniejszej aktywacji receptorów DA (Yokel and Wise, 1975; Di Chiara i Imperato, 1988; Bergman i in., 1990; Kehoe i in., 1996). Dlatego zmiany w każdym z tych czynników mogą leżeć u podstaw wpływu doświadczenia wiązania par na nagrodę AMPH. Poziomy uwolnionej DA były ściśle skorelowane z pozytywnymi subiektywnymi skutkami nadużywania narkotyków (Volkow i wsp., 1999; Drevets i in., 2001; Leyton, 2010) i uwalnianie DA wywołane środkami psychostymulacyjnymi NAcc zostało zmienione przez doświadczenia społeczne w niektórych przypadkach [np. izolacja we wczesnym okresie życia (Kehoe i in., 1996; Kosten i in., 2005)], ale nie inne [np. wzbogacone budownictwo socjalne (Bardo i in., 1995)]. W niniejszym badaniu leczenie AMPH zwiększyło poziom pozakomórkowej DA w NAcc, co jest zgodne z wcześniejszymi doniesieniami (Di Chiara i Imperato, 1988; Curtis i Wang, 2007; McKittrick i Abercrombie, 2007). Ponadto AMPH wywołał natychmiastowy i długotrwały spadek poziomu zewnątrzkomórkowego DOPAC w obu grupach. Odkrycie to jest zgodne z wynikami poprzednich badań i znaną rolą AMPH w hamowaniu oksydazy monoaminowej - enzymu zaangażowanego w degradację DA (Zielony i el Hait, 1978; Jones i in., 1998; Curtis i Wang, 2007). W obu przypadkach wielkość i czasowy wzorzec zmian pozakomórkowych w NAcc były porównywalne między samcami SN i PB, co sugeruje, że jest mało prawdopodobne, aby efekty wiązania par na nagrodę AMPH były związane z uwalnianiem lub metabolizmem DA NAcc.
Chociaż doświadczenie wiązania par nie miało wpływu na uwalnianie lub metabolizm DA indukowane przez AMPH, miało to wpływ na działanie AMPH na wiązanie receptora DA NAcc (tj. ekspresję i/lub powinowactwo receptora DA). Na przykład, zgodnie z poprzednimi badaniami, leczenie AMPH zwiększyło ekspresję NAcc D1R u mężczyzn SN (Liu i wsp., 2010) i doświadczenie wiązania par również podniosło wiązanie NAcc D1R (Aragona i in., 2006). Jednak wpływ AMPH na wiązanie NAcc D1R był odwrócony u samców PB w porównaniu z samcami SN, ponieważ samce PB wykazywały znaczny spadek wiązania D1R po leczeniu AMPH. Nie stwierdzono różnic między grupami w wiązaniu NAcc D2R ani w bieżących, ani w poprzednich badaniach (Aragona i in., 2006; Liu i wsp., 2010). Łącznie dane te pokazują, że doświadczenie wiązania par i ekspozycja na AMPH prowadzi do zmian specyficznych dla D1R w NAcc samców SN. Ponadto, ponieważ AMPH wpływało odwrotnie na wiązanie NAcc D1R u norników SN i PB, nasze dane wskazują, że więzi społeczne mogą być ważnym czynnikiem pośredniczącym we wpływie AMPH na mezolimbiczny układ DA. Odpowiednio, stwierdzono, że kokaina indukuje silną pozytywną zmianę sygnału zależnego od poziomu natlenienia krwi (BOLD) w całym systemie mezolimbicznej DA u dziewiczych samic, ale w dużej mierze negatywną odpowiedź BOLD u matek karmiących, mierzoną funkcjonalnym rezonansem magnetycznym (Ferris i in., 2005), tym samym wskazując dalej, że doświadczenia społeczne/seksualne mogą odgrywać ważną rolę w neurobiologicznej odpowiedzi na nadużywane narkotyki. Zmiany w wiązaniu receptora, takie jak te opisane powyżej, mogą mieć głęboki wpływ na zachowanie, ponieważ modyfikują reakcję mózgu na uwolnione neuroprzekaźniki. Na przykład u samców SN, indukowany przez AMPH wzrost wiązania D1R może odgrywać ważną rolę podczas kondycjonowania AMPH, ponieważ blokada D1R w NAcc, ale nie D2R, hamowała CPP indukowaną przez AMPH – odkrycie zgodne z tymi u innych gatunków (Baker i in., 1998; Pitchers i wsp., 2010). W konsekwencji, u samców PB, zmniejszone wiązanie NAcc D1R może leżeć u podstaw braku CPP indukowanego przez AMPH, ponieważ aktywacja NAcc D1R podczas kondycjonowania AMPH umożliwiła CPP indukowaną przez AMPH u mężczyzn PB. Łącznie, dane te wskazują, że indukowane przez AMPH spadki ekspresji/powinowactwa NAcc D1R mogą leżeć u podstaw wpływu doświadczenia wiązania par na nagrodę AMPH.
Warto wspomnieć o dwóch interesujących podobieństwach między naszymi odkryciami a wynikami innych badań, które badały związek między nadużywaniem narkotyków a zachowaniami społecznymi. Po pierwsze, wiązanie par i wielokrotna ekspozycja na AMPH niezależnie od siebie powodowały podobne zmiany w mezolimbicznym systemie DA samców norników preriowych [tj. wzmocnione wiązanie D1R (Aragona i in., 2006) i wyrażenie (Liu i wsp., 2010) w NAcc]. Podobnie, doświadczenie seksualne i wielokrotna ekspozycja na psychostymulanty zwiększały gęstość kolców dendrytycznych na średnich neuronach kolczastych w powłoce NAcc u szczurów (Robinson i Kolb, 1999; Pitchers i wsp., 2010). Ponadto, ssanie szczeniąt w matkach karmiących i podawanie kokainy u dziewiczych samic wywołało podobne wzorce dodatniej aktywacji BOLD w układzie mezokortykolimbicznym (Ferris i in., 2005). Łącznie dane te potwierdzają pogląd, że nadużywane narkotyki uzurpują sobie mechanizmy nerwowe i obwody, które pośredniczą w zachowaniach adaptacyjnych (Panksepp i in., 2002). Po drugie, nasze dane sugerują, że zróżnicowane reakcje neurobiologiczne na AMPH u samców SN i PB mogą leżeć u podstaw różnic między grupami w zachowaniu wywołanym AMPH. Podobnie, karmiące piersią szczury wykazywały tłumienie aktywności w mezokortykolimbicznym systemie DA w odpowiedzi na kokainę (w przeciwieństwie do wzrostu aktywności odnotowanego u dziewiczych samic) (Ferris i in., 2005) i zmniejszenie samopodawania kokainy (Hecht i in., 1999), dalej wspierając pogląd, że zmiany fizjologiczne związane z doświadczeniem społecznym mogą osłabiać satysfakcjonujące właściwości narkotyków.
Obecność silnych więzi społecznych w wieku dorosłym może zmniejszyć podatność na nadużywanie narkotyków (Kosten i in., 1987). Jednak mechanizmy neuronalne leżące u podstaw tego zjawiska behawioralnego są stosunkowo nieznane, być może z powodu braku odpowiedniego modelu zwierzęcego. W niniejszym badaniu ustaliliśmy nornicę preriową jako model zwierzęcy, aby zbadać mechanizmy neuronalne leżące u podstaw ochronnego wpływu więzi społecznych dorosłych na podatność na nadużywanie narkotyków. Nasze odkrycia wskazują, że doświadczenie wiązania par zmniejsza nagradzające właściwości AMPH i że mezolimbiczny system DA, szczególnie neurotransmisja DA w NAcc, pośredniczy w tym efekcie. Te wyniki, wraz z wynikami naszego poprzedniego badania (Liu i wsp., 2010), ustanowienie podstaw dla przyszłych badań mechanizmów neuronalnych leżących u podstaw wzajemnego związku między używaniem / nadużywaniem narkotyków a więzami społecznymi, co może ostatecznie zapewnić ważny wgląd w zapobieganie lub leczenie nadużywania narkotyków.